Biosynthesis ng kolesterol at biochemistry nito - Diabetes
Walang pag-aalinlangan, ang kolesterol ay ang pinakamahusay na kilalang lipid sa pangkalahatang publiko; ito ay kilalang-kilala dahil sa mataas na ugnayan sa pagitan ng mataas na kolesterol ng dugo at ang dalas ng mga sakit sa cardiovascular ng tao. Ang mas kaunting pansin ay nabayaran sa mahalagang papel ng kolesterol bilang isang bahagi ng mga lamad ng cell at bilang isang hudyat sa mga hormone ng steroid at mga acid ng apdo. Ang kolesterol ay kinakailangan para sa maraming mga hayop, kabilang ang mga tao, ngunit ang pagkakaroon nito sa pagkain ng mammal ay opsyonal - ang mga cell ng katawan mismo ay maaaring synthesize ito mula sa mga simpleng precursors.
Ang istraktura ng 27-carbon compound na ito ay nagmumungkahi ng isang komplikadong landas para sa biosynthesis nito, ngunit ang lahat ng mga carbon atoms na ito ay ibinibigay ng isang solong precursor - acetate. Mga bloke ng Isoprene - ang pinakamahalagang tagapamagitan mula sa acetate hanggang sa kolesterol, ang mga ito ay ang mga nauna sa maraming likas na lipid, at ang mga mekanismo sa pamamagitan ng mga bloke ng isoprene ay polimerisado ay katulad sa lahat ng mga metabolic pathway.
Nagsisimula kami sa pamamagitan ng pagsusuri sa mga pangunahing yugto sa landas ng biosynthesis ng kolesterol mula sa acetate, pagkatapos ay talakayin ang transportasyon ng kolesterol sa pamamagitan ng daloy ng dugo, ang pagsipsip ng mga cell, ang normal na regulasyon ng synthesis ng kolesterol, at regulasyon sa mga kaso ng impaired pagsipsip o transportasyon. Pagkatapos ay titingnan namin ang iba pang mga sangkap na nagmumula sa kolesterol, tulad ng mga acid ng bile at mga hormone ng steroid. Sa wakas, ang isang paglalarawan ng mga biosynthetic na mga landas para sa pagbuo ng maraming mga compound - mga derivatives ng isoprene blocks, kung saan mayroong mga karaniwang mga unang yugto na may synt synthesis ng kolesterol, ay ilalarawan ang pambihirang kakayahang magamit ng pagkakaugnay ng isoprenoid sa biosynthesis.
Ang kolesterol ay ginawa mula sa acetyl-CoA sa apat na yugto
Ang kolesterol, tulad ng mahabang chain fatty fatty, ay ginawa mula sa acetyl-CoA, ngunit ang pattern ng pagpupulong ay ganap na naiiba. Sa mga unang eksperimento, ang acetate na may label na may 14 C alinman sa methyl o carboxyl carbon atom ay idinagdag sa feed ng hayop. Batay sa pamamahagi ng label sa kolesterol na nakahiwalay sa dalawang pangkat ng mga hayop (Larawan 21-32), inilarawan ang mga yugto ng enzymatic ng biosynthesis ng kolesterol.
Fig. 21-32. Pinagmulan ng carbon atoms ng kolesterol. Nakilala sa panahon ng mga eksperimento gamit ang radioactive acetate na may label na may methyl carbon (itim) o carboxyl carbon (pula). Sa nakakabuo na istraktura, ang mga singsing ay minarkahan ng mga titik A hanggang D.

Ang synthesis ay naganap sa apat na yugto, tulad ng ipinapakita sa Fig. 21-33: (1) ang paghalay ng tatlong residu ng acetate upang mabuo ang isang anim na carbon intermediate ng mevalonate, (2) ang pagbabagong-anyo ng mevalonate upang maisaaktibo ang mga bloke ng isoprene, (3) ang polimerisasyon ng anim na limang yunit ng isterrene na carbon upang mabuo ang isang 30-carbon linear squalene, (4) pag-ikot ng squalene upang mabuo apat na singsing ng nucleus ng steroid, na sinundan ng isang serye ng mga pagbabago (oksihenasyon, pag-alis o paglipat ng mga grupo ng methyl) na may pagbuo ng kolesterol.
Fig. 21-33. Pangkalahatang larawan ng biosynthesis ng kolesterol. Apat na yugto ng synthesis ang tinalakay sa teksto. Ang mga bloke ng Isoprene sa squalene ay minarkahan ng mga pulang linya.

Stage (1). Sintesis ng mevalonate mula sa acetate. Ang unang yugto ng biosynthesis ng kolesterol ay humahantong sa pagbuo ng isang intermediate na produkto mevalonate (Larawan 21-34). Ang dalawang acetyl CoA Molekyul ay nagbibigay sa pagbibigay ng acetoacetyl CoA, na nakakabit sa ikatlong acetyl CoA molekula upang mabuo ang isang anim na carbon compound β-hydroxy-β-methylglutaryl-CoA (HM G -CoA). Ang dalawang unang reaksyon na ito ay catalyzed thiolase at NM G -CoA synthase, ayon sa pagkakabanggit. Cytosolic Synthase ng NM G-CoA Ang metabolic pathway na ito ay naiiba sa mitochondrial isoenzyme, na catalyzes ang synthesis ng NM G -CoA sa panahon ng pagbuo ng ketone body (tingnan ang Fig. 17-18).
Fig. 21-34. Ang pagbuo ng mevalonate mula sa acetyl-CoA. Ang mapagkukunan ng C-1 at C-2 mevalonate mula sa acetyl-CoA ay naka-highlight sa rosas.

Ang ikatlong reaksyon ay nililimitahan ang bilis ng buong proseso. Sa loob nito, ang NM G -CoA ay nabawasan sa mevalonate, kung saan ang bawat isa sa dalawang mga molekulang NА D PH ay nagbibigay ng dalawang elektron. HMG-CoA reductase - integral lamad protina ng makinis na ER, nagsisilbi ito, tulad ng makikita natin sa ibang pagkakataon, bilang pangunahing punto ng regulasyon ng metabolic pathway ng pagbuo ng kolesterol.
Stage (2). Ang pag-convert ng mevalonate sa dalawang aktibo na isoprene. Sa susunod na yugto ng synthesis ng kolesterol, tatlong grupo ng pospeyt ang inilipat mula sa mga molekula ng ATP hanggang mevalonate (Fig. 21-35). Ang pospeyt na nakagapos sa pangkat ng hydroxyl sa C-3 mevalonate sa intermediate 3-phospho-5-pyrophosphomevalonate ay isang mahusay na pag-iiwan ng grupo, sa susunod na hakbang pareho ng mga pospeyt na ito at ang katabing pangkat ng carboxyl leave, na bumubuo ng isang dobleng bono sa limang produktong carbon ∆ 3 -isopentenyl pyrophosphate. Ito ang una sa dalawang aktibong isoprenes - ang pangunahing mga kalahok sa synthesis ng kolesterol. Ang isomerization ng Δ 3 -isopentenylpyrophosphate ay nagbibigay ng isang pangalawang naaktibo na isoprene dimethylallyl pyrophosphate. Ang synthesis ng isopentenyl pyrophosphate sa cytoplasm ng mga cell cells ay nangyayari ayon sa landas na inilarawan dito. Gayunpaman, ang mga chloroplast ng halaman at maraming bakterya ay gumagamit ng isang landas na independiyenteng ng mevalonate. Ang alternatibong ruta na ito ay hindi matatagpuan sa mga hayop, kaya kaakit-akit kapag lumilikha ng mga bagong antibiotics.
Fig. 21-35. Ang conversion ng mevalonate sa na-activate na mga bloke ng isoprene Ang anim na aktibo na yunit ay pinagsama upang makabuo ng squalene (tingnan ang Larawan 21-36). Ang mga umaalis na grupo ng 3-phospho-5-pyrophosphomevalonate ay naka-highlight sa rosas. Sa square bracket ay isang hypothetical intermediate.

Stage (3). Ang kondensasyon ng anim na aktibo na yunit ng isoprene upang makabuo ng squalene. Ang Isopentenyl pyrophosphate at dimethylallyl pyrophosphate ngayon ay sumasailalim sa kondensasyon ng ulo-sa-buntot, kung saan ang isang pangkat na pyrophosphate ay gumagalaw at isang 10-carbon chain form - geranyl pyrophosphate (Larawan 21-36). (Ang Pyrophosphate ay nakakabit sa ulo.) Ang Geranyl pyrophosphate ay sumasailalim sa sumusunod na kondensasyon ng ulo-sa-buntot na may isopentenyl pyrophosphate, at isang 15-carbon intermediate form farnesyl pyrophosphate. Sa wakas, ang dalawang molekula ng farnesyl pyrophosphate ay pinagsama ang "ulo sa ulo", ang parehong mga grupo ng pospeyt ay tinanggal - nabuo squalene.
Fig. 21-36. Pagbubuo ng squalene. Ang isang istraktura ng squalene na naglalaman ng 30 carbon atoms ay nangyayari sa sunud-sunod na condensations na isinaaktibo ng mga bloke ng isoprene (five-carbon).

Ang mga karaniwang pangalan para sa mga tagapamagitan na ito ay nagmula sa mga pangalan ng mga mapagkukunan kung saan sila ay unang nakahiwalay. Ang Geraniol, isang bahagi ng langis ng rosas, ay may lasa ng geranium, at farnesol, na matatagpuan sa mga kulay ng acacia farnesa, ay may liryo ng aroma ng lambak. Maraming mga likas na amoy ng halaman ay nabibilang sa mga compound na binuo mula sa mga bloke ng isoprene. Ang squalene, una na nakahiwalay sa atay ng pating (Squalus species), ay binubuo ng 30 carbon atoms: 24 atoms sa pangunahing kadena at anim na atoms sa mga substituents ng metal.
Stage (4). Ang pagbabagong-anyo ng squalene sa apat na singsing ng isang steroid na inti. Sa fig. 21-37 malinaw na nakikita na ang istruktura ng chain ng squalene, at sterols - siklik. Ang lahat ng mga sterol ay may apat na condensing singsing na bumubuo ng steroid na nucleus, at lahat ng mga ito ay mga alkohol na may pangkat na hydroxyl sa C-3 atom, samakatuwid ang Ingles na pangalan na sterol. Sa ilalim ng pagkilos squalene m soaxygenase isang oxygen atom mula sa O ay idinagdag sa dulo ng chain ng squalene 2 at isang epoxide ay nabuo. Ang enzyme na ito ay isa pang mixed-function na oxidase (idagdag. 21-1), binabawasan ng NADPH ang isa pang atom na oxygen mula sa O 2 sa H2 O. Mga Doble ng Ikalawang Produkto squalene-2,3-epoxide inayos upang ang isang kapansin-pansin na reaksyon ay maaaring maging isang kadena ng squalene epoxide sa isang istraktura ng paikot. Sa mga cell ng hayop, ang pagbibisikleta na ito ay humahantong sa lanosterol na naglalaman ng apat na singsing na katangian ng steroid nucleus. Bilang isang resulta, angosteroster ay na-convert sa kolesterol sa pamamagitan ng isang serye ng humigit-kumulang na 20 reaksyon, na kasama ang paglipat ng ilang mga grupo ng metal at pagtanggal ng iba. Ang paglalarawan ng kamangha-manghang landas ng biosynthesis, isa sa pinakamahirap sa mga kilala, ay ginawa nina Conrad Bloch, Theodore Linen, John Cornfort at George Popiak sa huling bahagi ng 1950s.
Fig. 21-37. Ang pagsasara ng singsing ay lumiliko ang linear squalene sa isang condensed steroid core. Ang unang yugto ay catalyzed ng isang oxidase na may isang halo-halong pag-andar (mtooxygenase), na ang cosubstrate ay N AD PH. Ang produkto ay isang epoxide, na sa susunod na yugto ay nag-cyclize upang makabuo ng isang core ng steroid. Ang pangwakas na produkto ng mga reaksyon na ito sa mga selula ng hayop ay kolesterol; sa iba pang mga organismo ay naiiba ang bahagyang naiiba mula dito.


Ang kolesterol ay isang sterol na katangian ng mga selula ng hayop, halaman, fungi at protists ay gumagawa ng iba pang mga katulad na mga sterol.
Ginagamit nila ang parehong ruta ng synthesis sa squalene-2,3-epoxide, ngunit pagkatapos ay ang mga landas ay lumihis nang bahagya, at iba pang mga sterols ay nabuo, tulad ng sigmasterol sa maraming mga halaman at ergosterol sa fungi (Fig. 21-37).
Halimbawa 21-1 Mga Gastos ng Enerhiya para sa Sintesis ng Squalene
Ano ang mga gastos sa enerhiya (na ipinahayag bilang mga molekula ng ATP) para sa synthesis ng isang molekula ng squalene?
Solusyon. Sa synthesis ng squalene mula sa acetyl-CoA, ang ATP ay ginugol lamang sa entablado kapag ang mevalonate ay na-convert sa isang activated isoprene squalene precursor. Ang anim na aktibong isoprene molekula ay kinakailangan upang makabuo ng isang molekula molekula, at tatlong molekula ng ATP ay kinakailangan upang makabuo ng bawat aktibong molekula. Sa kabuuan, 18 na molekula ng ATP ang ginugol sa synthesis ng isang squalene molekula.
Mga Compound ng kolesterol sa katawan
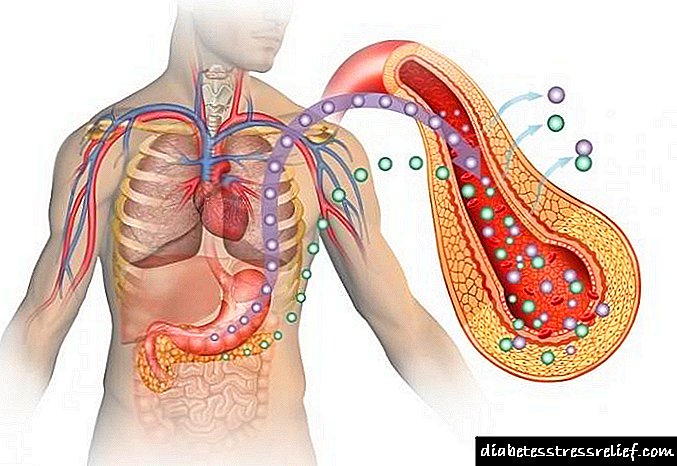
Sa mga vertebrates, ang malaking halaga ng kolesterol ay synthesized sa atay. Ang ilan sa mga kolesterol na synthesized doon ay nakasama sa mga lamad ng hepatocytes, ngunit higit sa lahat ito ay nai-export sa isa sa tatlong mga form nito: biliary (apdo) kolesterol, apdo acid o kolesterol esters. Mga acid acid at ang kanilang mga asing asin ay hydrophilic derivatives ng kolesterol, na synthesized sa atay at nag-ambag sa pagtunaw ng mga lipid (tingnan ang Fig. 17-1). Mga Ester ng kolesterol nabuo sa atay sa pamamagitan ng pagkilos acyl-CoA-cholesterol-acyltransferase (ACAT). Ang enzyme na ito ay catalyzes ang paglipat ng isang natitirang fatty acid mula sa coenzyme A hanggang sa hydroxyl group ng kolesterol (Fig. 21-38), na nagiging kolesterol ang anyo ng kolesterol. Ang mga esters ng kolesterol sa mga lihim na lipoprotein na mga particle ay dinadala sa iba pang mga tisyu gamit ang kolesterol o nakaimbak sa atay.
Fig. 21-38. Sintesis ng mga kolesterol ester. Ang Etherification ay gumagawa ng kolesterol na isang mas hydrophobic form para sa imbakan at transportasyon.

Ang kolesterol ay kinakailangan para sa lahat ng mga tisyu ng isang lumalagong organismo ng hayop para sa synthesis ng mga lamad, at ilang mga organo (halimbawa, ang mga adrenal glandula at glandula ng sex) ay gumagamit ng kolesterol bilang isang paunang-una sa mga hormone ng steroid (tatalakayin ito sa ibaba). Ang kolesterol ay isa ring pasiuna sa bitamina D (tingnan ang Larawan 10-20, v. 1).
Ang kolesterol at iba pang mga lipid ay nagdadala ng plasma lipoproteins
Ang mga ester sa kolesterol at kolesterol, tulad ng mga triacylglycerol at phospholipids, ay praktikal na hindi malulutas sa tubig, gayunpaman, dapat silang lumipat mula sa tisyu kung saan sila ay synthesized sa mga tisyu kung saan sila ay maiimbak o maubos. Ang mga ito ay dinadala ng daloy ng dugo sa anyo ng dugo plasma lipoproteins - macromolecular complexes ng mga tiyak na protina ng carrier (apolipoproteins) na may mga pospolipid, kolesterol, kolesterol esters at triacylglycerols na naroroon sa mga kumplikadong ito sa iba't ibang mga kumbinasyon.
Ang Apolipoproteins ("apo" ay tumutukoy sa protina ng lipid-free na sarili) pagsamahin sa mga lipid upang makabuo ng ilang mga praksiyon ng mga particle ng lipoprotein - mga spherical complex na may hydrophobic lipids sa gitna at hydrophilic amino acid chain sa ibabaw (Fig. 21-39, a). Sa iba't ibang mga kumbinasyon ng mga lipid at protina, ang mga partikulo ng iba't ibang mga density ay nabuo - mula sa mga chylomicrons hanggang sa mataas na density lipoproteins. Ang mga particle na ito ay maaaring paghiwalayin ng ultracentrifugation (Talahanayan 21-1) at biswal na sinusunod gamit ang electros microscopy (Larawan 21-39, b). Ang bawat bahagi ng lipoproteins ay gumaganap ng isang tukoy na pag-andar, na natutukoy ng lugar ng synthesis, lipid na komposisyon at nilalaman ng apolipoprotein. Hindi bababa sa 10 iba't ibang mga apolipoproteins ay natagpuan sa plasma ng dugo ng tao (Talahanayan 21-2), na nag-iiba sa laki, reaksyon na may tiyak na mga antibodies, at ang katangian na pamamahagi sa iba't ibang klase ng lipoproteins. Ang mga sangkap na protina na ito ay nagsisilbing senyales ng mga sangkap na nagdidirekta ng mga lipoprotein sa mga tukoy na tisyu o pag-activate ng mga enzyme na kumikilos sa lipoproteins.
Talahanayan 21-1. Human plasma lipoproteins
Komposisyon (maliit na bahagi,%)
r = 513,000). Ang isang maliit na butil ng LDL ay naglalaman ng isang core ng humigit-kumulang na 1,500 molekula ng mga estersang kolesterol, sa paligid ng pangunahing mayroong isang shell ng 500 na mga molekula ng kolesterol, 800 na mga molekula ng phospholipid at isang molekula ng apoB-100. b - apat na klase ng lipoproteins, na nakikita ng isang mikroskopyo ng elektron (pagkatapos ng paghahayag ng negatibo). Clockwise, simula sa itaas na kaliwang pigura: chylomicrons - na may diameter na 50 hanggang 200 nm, PL O NP - mula 28 hanggang 70 nm, HDL - mula 8 hanggang 11 nm, at LDL - mula 20 hanggang 55 nm. Ang mga katangian ng lipoproteins ay ibinibigay sa talahanayan. 21-2.

Chylomicrons, tinukoy sa Sec. 17, ilipat ang mga triacylglycerol ng pagkain mula sa bituka sa iba pang mga tisyu. Ito ang pinakamalaking lipoproteins, mayroon silang pinakamababang density at ang pinakamataas na kamag-anak na nilalaman ng triacylglycerol (tingnan ang Fig. 17-2). Ang mga chylomicrons ay synthesized sa ER ng mga epithelial cells na naglalagay ng maliit na bituka, pagkatapos ay ilipat sa pamamagitan ng lymphatic system at ipasok ang daloy ng dugo sa pamamagitan ng kaliwang subclavian vein. Ang Chylomicron apolipoproteins ay naglalaman ng apoB-48 (natatangi para sa klase ng lipoproteins), apoE at apoC-II (Talahanayan 21-2). Aktibo ng AroC-II ang lipoprotein lipase sa mga capillary ng adipose tissue, puso, kalamnan ng kalansay at paggagatas ng glandula ng mammary, na tinitiyak ang daloy ng mga libreng fatty acid sa mga tisyu. Sa gayon, ang mga chylomicrons ay naglilipat ng mga fatty fatty acid sa mga tisyu, kung saan sila ay maubos o maiimbak bilang gasolina (Larawan 21-40). Ang mga residue ng Chylomicron (higit sa lahat ay napalaya mula sa triacylglycerols, ngunit naglalaman pa rin ng kolesterol, apoE at apoB-48) ay dinala ng daloy ng dugo patungo sa atay. Sa atay, ang mga receptor ay nagbubuklod sa apoE na nakapaloob sa mga residu ng chylomicron at pag-mediate ang kanilang pagsipsip sa pamamagitan ng endocytosis. Sa mga hepatocytes, inilalabas ng mga residue na ito ang kolesterol na naglalaman ng mga ito at nawasak sa mga lysosome.
Talahanayan 21-2. Human plasma lipoprotein apolipoproteins
Function (kung kilala)
Aktibo ang L CAT, nakikipag-ugnay sa transporter ng ABC
Nagpapakita ng L CAT
Aktibo ang L CAT, kolesterol transport / clearance
Binds sa LDL receptor
Mga Chylomicrons, VLDL, HDL
Mga Chylomicrons, VLDL, HDL
Mga Chylomicrons, VLDL, HDL
Nagsisimula ang clearance ng VLDL at residue ng chylomicron
Kung ang pagkain ay naglalaman ng mas maraming mga fatty acid kaysa sa kasalukuyan ay maaaring magamit bilang gasolina, nagiging mga triacylglycerol sa atay, na bumubuo ng isang bahagi na may mga tiyak na apolipoproteins napakababang density lipoproteins (VLDL). Ang sobrang karbohidrat sa atay ay maaari ring ma-convert sa triacylglycerols at ma-export bilang VLDL (Larawan 21-40, a).Bilang karagdagan sa mga triacylglycerol, ang maliit na bahagi ng VLDL ay naglalaman ng isang tiyak na halaga ng mga ester ng kolesterol at kolesterol, pati na rin apoB-100, apoC-1, apoC-II, apoC III at apoE (Talahanayan 21-2). Ang mga lipoproteins na ito ay dinadala din ng dugo mula sa atay patungo sa kalamnan at adipose tissue, kung saan, pagkatapos ng lipoprotein lipase ay naisaaktibo ng apo-C II, ang mga libreng fatty acid ay pinalaya mula sa triacylglycerols ng VDL na bahagi. Ang mga Adipocytes ay nakakuha ng mga libreng fatty acid, muli itong ibinalik sa triacylglycerols, na nakaimbak sa mga cells na ito sa anyo ng mga inclusions ng lipid (patak), myocytes, sa kabaligtaran, agad na oxidize ang mga fatty acid upang makabuo ng enerhiya. Karamihan sa mga residue ng VLDL ay tinanggal mula sa sirkulasyon ng mga hepatocytes. Ang kanilang pagsipsip, na katulad ng pagsipsip ng chylomicrons, ay pinagsama ng mga receptor at nakasalalay sa pagkakaroon ng apoE sa mga residue ng VLDL (bilang karagdagan. 21-2 ang kaugnayan sa pagitan ng apoE at Alzheimer na sakit ay inilarawan).
Fig. 21-40. Ang mga lipoproteins at transportasyon ng lipid, at - mga lipid ay dinadala ng daloy ng dugo sa anyo ng mga lipoproteins, na pinagsama sa ilang mga praksyon na may iba't ibang mga pag-andar at iba't ibang komposisyon ng mga protina at lipid (tab. 21-1, 21-2) at tumutugma sa density ng mga fraction na ito. Ang mga lipid ng pagkain ay pinagsama-sama sa mga chylomicrons, karamihan sa mga triacylglycerol na nilalaman nito ay pinakawalan ng lipoprotein lipase sa adipose at kalamnan tissue sa mga capillary. Ang mga residue ng Chylomicron (naglalaman ng pangunahing protina at kolesterol) ay nakuha ng mga hepatocytes. Ang mga endogenous lipids at kolesterol mula sa atay ay inihatid sa adipose at kalamnan tissue sa anyo ng VLDL. Ang pagpapalabas ng mga lipid mula sa VLDL (kasama ang pagkawala ng ilang mga apolipoproteins) ay unti-unting na-convert ang VLDLP sa LDL, na naghahatid ng kolesterol sa mga tisyu ng extrahepatic o ibabalik ito sa atay. Kinukuha ng atay ang mga labi ng VLDL, LDL at ang mga labi ng chylomicrons sa pamamagitan ng receptor-mediated endocytosis. Ang sobrang kolesterol sa mga tisyu ng extrahepatic ay dinadala pabalik sa atay sa anyo ng LDL. Sa atay, ang bahagi ng kolesterol ay nagiging mga asin ng apdo. b - mga sample ng plasma ng dugo na nakuha pagkatapos ng gutom (kaliwa) at pagkatapos kumain ng pagkain na may mataas na nilalaman ng taba (kanan). Ang mga chylomicrons na nabuo sa pamamagitan ng pagkain ng mga mataba na pagkain ay nagbibigay sa plasma ng isang panlabas na pagkakahawig sa gatas.

Sa pagkawala ng mga triacylglycerols, ang isang bahagi ng VLDL ay na-convert sa mga residue ng VLDL, na tinatawag ding intermediate density lipoproteins (VLDL), karagdagang pag-alis ng triacylglycerols mula sa VLDL ay nagbibigay mababang density lipoproteins (LDL) (tab. 21-1). Ang maliit na bahagi ng LDL, na mayaman sa kolesterol at kolesterol esters, at naglalaman din ng apoB-100, naglilipat ng kolesterol sa mga tisyu ng extrahepatic na nagdadala ng mga tukoy na receptor na kinikilala ang apoB-100 sa kanilang mga lamad ng plasma. Ang mga receptor na ito ay nagpapagitna sa pag-aalsa ng mga estersang kolesterol at kolesterol (tulad ng inilarawan sa ibaba).
Pagdaragdag 21-2.Alamin ng mga alelasyon ng ApoE ang saklaw ng sakit na Alzheimer
Sa populasyon ng tao, mayroong tatlong kilalang mga variant (tatlong alleles) ng gene na naka-encode ng apolipoprotein E. Ng apoel alleles, ang alleles ng APOEZ ay ang pinaka-karaniwan sa mga tao (tungkol sa 78%), ang APOE4 at APOE2 alleles ay 15 at 7%, ayon sa pagkakabanggit. Ang APOE4 allele ay lalo na katangian ng mga taong may sakit na Alzheimer, at pinapayagan ang ugnayang ito na mahulaan ang paglitaw ng sakit na may mataas na posibilidad. Ang mga taong nagmana sa APOE4 ay may mataas na peligro sa pagbuo ng huli na Alzheimer's disease. Ang mga taong homozygous para sa APOE4 ay 16 na beses na mas malamang na magkaroon ng sakit, ang average na edad ng mga taong nagkakasakit ay mga 70 taong gulang. Para sa mga taong nagmana ng dalawang kopya ng AROEZ, sa kabaligtaran, ang average na edad ng sakit ng Alzheimer ay lumampas sa 90 taon.
Ang pang-molekular na batayan para sa ugnayan sa pagitan ng apoE4 at sakit ng Alzheimer ay hindi pa kilala. Bilang karagdagan, hindi pa malinaw kung paano maapektuhan ng apoE4 ang paglaki ng mga amyloid cords, na tila ang ugat ng sakit ng Alzheimer (tingnan ang Fig. 4-31, v. 1). Ang mga pagpapalagay ay nakatuon sa posibleng papel ng apoE sa pag-stabilize ng istraktura ng cytoskeleton ng mga neuron. Ang mga protina ng apoE2 at apoEZ ay nagbubuklod sa isang bilang ng mga protina na nauugnay sa microtubule ng mga neuron, habang ang apoE4 ay hindi nagbubuklod. Maaari itong mapabilis ang pagkamatay ng mga neuron. Anuman ang mekanismong ito ay maaaring maging, ang mga obserbasyong ito ay nagbibigay ng pag-asa para sa pagpapalawak ng aming pag-unawa sa mga biological function ng apolipoproteins.
Ang ika-apat na uri ng lipoproteins - Mataas na density lipoproteins (HDL), ang bahagi na ito ay nabuo sa atay at maliit na bituka sa anyo ng maliit na mga particle na mayaman sa protina na naglalaman ng medyo maliit na kolesterol at ganap na walang koleksyon ng mga esters ng kolesterol (Larawan 21-40). Ang maliit na bahagi ng HDL ay naglalaman ng apoA-I, apoC-I, apoC-II at iba pang mga apolipoproteins (Table 21-2), pati na rin lecithin-cholesterol-acyltransferase (LC AT), na catalyzes ang pagbuo ng mga kolesterol esters mula sa lecithin (phosphatidylcholine) at kolesterol (Larawan 21-41). Ang L CAT sa ibabaw ng mga bagong nabuo na mga particle ng HDL ay nagko-convert ng chylomicron kolesterol at phosphatidylcholine at VLDL na natitira sa mga esters ng kolesterol, na nagsisimula upang mabuo ang nucleus, na binago ang bagong nabuo na mga particle na HDO na hindi nabuo sa mga mature spherical HDL particle. Ang lipoprotein na mayaman sa kolesterol na ito ay pagkatapos ay ibabalik sa atay, kung saan ang kolesterol ay "pinalabas", ang ilan sa kolesterol na ito ay na-convert sa mga bile salt.
Fig. 21-41. Ang reaksyon ay nabalisa ng lecithin-cholesterol-acyltransferase (L CAT). Ang enzyme na ito ay naroroon sa ibabaw ng mga particle ng HDL at naisaaktibo ng apoA-1 (isang bahagi ng bahagi ng HDL). Ang mga esters ng kolesterol ay natipon sa loob ng mga bagong nabuo na mga particle ng HDL, na nagiging mga ito sa mature HDL.

Ang HDL ay maaaring masisipsip sa atay sa pamamagitan ng receptor-mediated endocytosis, ngunit hindi bababa sa ilan sa HDL kolesterol ay naihatid sa iba pang mga tisyu ng iba pang mga mekanismo. Ang mga particle ng HDL ay maaaring magbigkis sa mga SR - BI receptor protein sa plasma lamad ng mga selula ng atay at sa mga steroid na tisyu tulad ng mga adrenal glandula. Ang mga receptor na ito ay hindi namamagitan sa endocytosis, ngunit bahagyang at pumipili paglipat ng kolesterol at iba pang mga lipid ng HDL na bahagi sa cell. Ang "naubos na" HDL na bahagi pagkatapos ay muling pumasok sa daloy ng dugo, kung saan kasama dito ang mga bagong bahagi ng mga lipid mula sa mga chylomicrons at residue ng VLDL. Ang parehong HDL ay maaari ring makuha ang kolesterol na nakaimbak sa extrahepatic tisyu at ilipat ito sa atay sa pamamagitan ng baligtarin ang transportasyon ng kolesterol (Larawan 21-40). Sa isa sa mga reverse variant ng transportasyon, ang pakikipag-ugnay ng nagresultang HDL kasama ang mga receptor ng SR-BI sa mga cells na mayaman sa kolesterol ay nagsisimula ng passive pagsasabog ng kolesterol mula sa ibabaw ng cell sa mga HDL particle, na pagkatapos ay ilipat ang kolesterol pabalik sa atay. Sa isa pang variant ng reverse transport sa isang rich kolesterol cell, pagkatapos ng pag-alis ng HDL, nakikipag-ugnay ang apoA-I sa aktibong transporter, ABC protein. Ang ApoA-I (at siguro HDL) ay nasisipsip ng endocytosis, pagkatapos ay na-sikreto muli, na puno ng kolesterol, na dinadala sa atay.
Ang Protein ABC1 ay bahagi ng isang malaking pamilya ng mga nagdadala ng maraming gamot, ang mga carrier na ito ay tinatawag minsan na mga transportasyon ng ABC, dahil lahat sila ay naglalaman ng mga ATP-binding cassette (ATP - nagbubuklod na mga cassette), mayroon din silang dalawang mga domain ng transmembrane na may anim na transmembrane helice (tingnan ang kap. 11, v. 1). Ang mga protina na ito ay aktibong naglilipat ng maraming mga ions, amino acid, bitamina, steroid hormones at apdo sa pamamagitan ng mga lamad ng plasma. Ang isa pang kinatawan ng pamilyang ito ng mga carrier ay ang protina ng CFTR, na, na may cystic fibrosis, ay nasira (tingnan ang dagdag. 11-3, v. 1).
Ang mga ester ng kolesterol ay pumapasok sa cell sa pamamagitan ng endocytosis ng receptor-mediated
Ang bawat butil ng LDL sa daloy ng dugo ay naglalaman ng apoB-100, na kinikilala ng mga tiyak na protina ng receptor ng ibabaw -Mga receptor ng LDL sa lamad ng mga cell na kailangang makunan ng kolesterol. Ang pagbubuklod ng LDL sa receptor ng LDL ay nagsisimula sa endocytosis, dahil sa kung saan lumipat ang LDL at ang receptor nito sa cell sa loob ng endosome (Fig. 21-42). Ang endosome sa kalaunan ay sumasama sa lysosome, na naglalaman ng mga enzyme na nag-hydrolyze ng mga esters ng kolesterol, naglalabas ng kolesterol at fatty acid sa cytosol. Ang ApoB-100 mula sa LDL ay nagbabawas din upang makabuo ng mga amino acid na na-sikreto sa cytosol, ngunit iniiwasan ng receptor ng LDL at bumalik sa cell ibabaw upang lumahok muli sa pagkalinga sa LDL. Ang ApoB-100 ay naroroon din sa VLDL, ngunit ang domain ng pagtanggap na ito ay hindi nakagapos sa LDL receptor; ang pag-convert ng VLDLP sa LDL ay ginagawang ang domain ng pagtanggap-binding sa apoB-100 naa-access. Ang landas ng transportasyon ng kolesterol sa dugo at ang endorterosis na mediated na receptor na ito sa mga target na tisyu ay pinag-aralan nina Michael Brown at Joseph Goldstein.
Michael Brown at Joseph Goldstein

Fig. 21-42. Ang pagkuha ng kolesterol sa pamamagitan ng receptor-mediated endocytosis.

Ang kolesterol, na pumapasok sa mga cell sa ganitong paraan, ay maaaring isama sa mga lamad o muling tinukoy ng ACAT (Fig. 21-38) para sa pag-iimbak sa cytosol sa loob ng mga patak ng lipid. Kung mayroong sapat na kolesterol na magagamit sa maliit na bahagi ng dugo ng LDL, ang akumulasyon ng labis na intracellular kolesterol ay pinipigilan sa pamamagitan ng pagbawas ng rate ng synthesis nito.
Ang LDL receptor ay nagbubuklod din sa apoE at gumaganap ng isang mahalagang papel sa paggana ng mga chylomicrons at residue ng VLDL sa atay. Gayunpaman, kung ang mga receptor ng LDL ay hindi magagamit (tulad ng, halimbawa, sa isang pilay ng mouse na may isang nawawalang LDL receptor gene), ang mga residue ng VLDL at chylomicrons ay hinihigop pa rin ng atay, bagaman ang LDL ay hindi hinihigop. Ipinapahiwatig nito ang pagkakaroon ng isang sistema ng pantulong na pantulong para sa mga receptor-mediated endocytosis ng VLDL at residue ng chylomicron. Ang isa sa mga reserbang reserbang ay ang protina ng LRP (lipoprotein receptor - nauugnay na protina), na nauugnay sa mga receptor ng lipoprotein, na nagbubuklod sa apoE at isang bilang ng iba pang mga ligand.
Maraming mga antas ng regulasyon ng biosynthesis ng kolesterol
Ang synthesis ng kolesterol ay isang kumplikado at masiglang na proseso, kaya malinaw na ang katawan ay kapaki-pakinabang na magkaroon ng isang mekanismo para sa pag-regulate ng biosynthesis ng kolesterol, na pinapunan ang halaga nito bilang karagdagan sa kung ano ang dala ng pagkain. Sa mga mammal, ang produksiyon ng kolesterol ay kinokontrol ng intracellular concentration
kolesterol at hormones glucagon at insulin. Ang yugto ng pag-convert ng HMG - CoA sa mevalonate (Fig. 21-34) ay nililimitahan ang bilis sa metabolic pathway ng pagbuo ng kolesterol (ang pangunahing punto ng regulasyon). Ang reaksyon na ito ay catalyzed ng HMG - CoA reductase. Ang regulasyon bilang tugon sa mga pagbabago sa mga antas ng kolesterol ay pinagsama ng isang matikas na sistema ng regulasyon ng transcriptional para sa isang gene na naka-encode ng HMG - CoA reductase. Ang gen na ito, kasama ang higit sa 20 iba pang mga gen na naka-encode ng mga enzyme na kasangkot sa pagsipsip at synthesis ng kolesterol at hindi nabubuong mga fatty acid, ay kinokontrol ng isang maliit na pamilya ng mga protina na tinatawag na mga protina na nakikipag-ugnay sa sterol-regulatory element ng pagbuo ng protina (SREBP, elementong regolohikal na nagbubuklod ng mga protina) . Pagkatapos ng synthesis, ang mga protina na ito ay ipinakilala sa endoplasmic reticulum. Ang tanging natutunaw na amino-terminal SREBP domain function bilang isang activator ng transkripsyon gamit ang mga mekanismo na inilarawan sa Ch. 28 (v. 3). Gayunpaman, ang domain na ito ay walang pag-access sa nucleus at hindi maaaring lumahok sa pag-activate ng gene hangga't nananatili ito sa molekula ng SREBP. Upang maisaaktibo ang transkripsyon ng gene ng HMG - CoA reductase at iba pang mga gen, ang domain na transkripsyonal ay nahihiwalay mula sa natitirang SREBP sa pamamagitan ng pag-clear ng proteolytic. Kapag ang kolesterol ay mataas, ang mga protina ng SREBP ay hindi aktibo, na naayos sa isang ER sa isang kumplikado na may isa pang protina na tinatawag na SCAP (SREBP - cleavage activating protein) (Fig. 21-43). Ito ay ang SCAP na nagbubuklod sa kolesterol at isang bilang ng iba pang mga sterol, na kumikilos bilang isang sensor ng sterol. Kung ang antas ng sterol ay mataas, ang SCAP - SREBP complex ay maaaring makipag-ugnay sa isa pang protina, na pinapanatili ang buong kumplikado sa ER. Kapag ang antas ng mga sterols sa cell ay bumababa, ang pagbabago ng conformational sa SCAP ay humantong sa pagkawala ng aktibidad ng pagpapanatili, at ang SCAP - SREBP complex ay lumilipat sa loob ng mga vesicle sa Golgi complex. Sa Golgi complex, ang mga protina ng SREBP ay na-doble ng dalawang beses sa pamamagitan ng dalawang magkakaibang mga proteases, ang pangalawang cleavage na naglabas ng amino-terminal domain sa cytosol. Ang domain na ito ay gumagalaw sa nucleus at isinaaktibo ang transkripsyon ng mga target na gen. Ang amino-terminal SREBP protein domain ay may isang maikling kalahating buhay at mabilis na pinanghihinaan ng mga proteasome (tingnan ang Fig. 27-48, t. 3). Kapag ang antas ng sterol ay tumaas nang sapat, ang pagpapalabas ng proteolytic ng mga protina ng SR EBP na protina gamit ang amino terminus ay muling hinarangan, at ang pagbagsak ng proteasome ng mga umiiral na aktibong domain ay humahantong sa isang mabilis na pagsara ng mga target na gen.
Fig. 21-43. Pag-activate ng SR EBP. Ang mga protina ng SREB P na nakikipag-ugnay sa isang elemento na regulado ng sterol (berde na kulay), kaagad pagkatapos ng synthesis, ay ipinakilala sa ER, na bumubuo ng isang kumplikadong may S CAP (pulang kulay). (Ang N at C ay nagpapahiwatig ng mga dulo ng amine at carboxyl ng mga protina.) Sa S-CAP na nakatali na estado, ang mga protina ng SRE BP ay hindi aktibo. Kapag bumababa ang antas ng sterol, ang SR EBP-S CAP complex ay lumilipat sa Golgi complex, at ang mga protina ng SR EBP ay sunud-sunod na na-clear ng dalawang magkakaibang mga protease. Ang liberated amino acid terminal SR EBP protein domain ay lumilipat sa nucleus, kung saan pinapagana nito ang transkripsyon ng mga genes na kinokontrol ng sterol.

Ang synthesis ng kolesterol ay kinokontrol din ng maraming iba pang mga mekanismo (Larawan 21-44). Ang control sa hormonal ay pinapamagitan ng covalent modification ng NM G-CoA reductase. Ang enzyme na ito ay umiiral sa phosphorylated (hindi aktibo) at dephosphorylated (aktibo) na form. Pinasisigla ng Glucagon ang phosphorylation (hindi aktibo) ng enzyme, at ang insulin ay nagtataguyod ng dephosphorylation, na-activate ang enzyme at pinapaboran ang synthesis ng kolesterol. Ang mataas na intracellular na konsentrasyon ng kolesterol ay nag-activate ng ASAT, na pinatataas ang esterification ng kolesterol para sa pag-aalis. Sa wakas, ang mga mataas na antas ng cellular cholesterol ay pumipigil sa transkripsyon ng gene na nag-encode ng LDL receptor, na binabawasan ang paggawa ng receptor na ito at, dahil dito, ang pagtaas ng kolesterol mula sa dugo.
Fig. 21-44. Ang regulasyon ng mga antas ng kolesterol ay nagbibigay ng isang balanse sa pagitan ng synthesis at pagsipsip ng kolesterol mula sa pagkain. Pinapabilis ng Glucagon ang phosphorylation (hindi aktibo) ng redmase ng NM G-CoA, ang insulin ay nagtataguyod ng dephosphorylation (activation). X - hindi nakilalang mga metabolite ng kolesterol na pinasisigla ang proteolysis ng NM G -CoA reductase.

Ang hindi regular na kolesterol ay maaaring humantong sa malubhang sakit sa mga tao. Kapag ang kabuuang halaga ng synthesized kolesterol at kolesterol na nakuha mula sa pagkain ay lumampas sa halaga na kinakailangan para sa pagpupulong ng lamad, synthesis ng mga bile salts at steroid, maaaring maiugnay ang mga pathological na akumulasyon ng kolesterol sa mga daluyan ng dugo (atherosclerotic plaques), na humahantong sa kanilang pagbara (atherosclerosis). Sa mga bansang industriyalisado, ito ay pagkabigo sa puso dahil sa hadlang sa coronary arteries na ang pangunahing sanhi ng dami ng namamatay. Ang pag-unlad ng atherosclerosis ay nauugnay sa isang mataas na antas ng kolesterol ng dugo at lalo na sa isang mataas na antas ng kolesterol na inilipat na may bahagi ng LDL, at isang mataas na antas ng dugo ng HDL, sa kabaligtaran, ay mainam na nakakaapekto sa estado ng mga daluyan ng dugo.
Sa namamana na hypercholesterolemia (isang genetic defect), ang antas ng kolesterol sa dugo ay napakataas - malubhang atherosclerosis ay nabuo sa mga taong ito na nasa pagkabata. Dahil sa isang may sira na LDL receptor, isang hindi sapat na pagtanggap-mediated na paggamit ng LDL kolesterol ay nangyayari. Bilang isang resulta, ang kolesterol ay hindi tinanggal mula sa daloy ng dugo, naipon ito at nag-aambag sa pagbuo ng mga atherosclerotic plaques. Ang synthesis ng endogenous kolesterol ay nagpapatuloy sa kabila ng labis na kolesterol sa dugo, dahil ang extracellular kolesterol ay hindi maaaring makapasok sa cell upang ayusin ang intracellular synthesis (Fig. 21 -44).Para sa paggamot ng mga pasyente na may namamana na hypercholesterolemia at iba pang mga sakit na nauugnay sa mataas na kolum ng suwero, ginagamit ang mga klase ng statin. Ang ilan sa mga ito ay nakuha mula sa mga likas na mapagkukunan, habang ang iba ay synthesized ng industriya ng parmasyutiko. Ang mga statins ay katulad ng mevalonate (magdagdag. 21-3) at mga mapagkumpitensyang inhibitor ng NMS-CoA reductase.
Pagdaragdag 21-3. MEDICINE. Ang lipid hypothesis at ang paglikha ng mga statins
Ang sakit sa coronary heart (CHD) ay ang pangunahing sanhi ng dami ng namamatay sa mga binuo bansa. Ang pagdidikit ng mga coronary artery na nagdadala ng dugo sa puso ay nangyayari bilang isang resulta ng pagbuo ng mga matitipid na deposito na tinatawag na atherosclerotic plaques; ang mga plake na ito ay naglalaman ng kolesterol, fibrillar protein, calcium, platelet clots, at cell fragment. Sa siglo XX. Nagkaroon ng aktibong debate tungkol sa ugnayan sa pagitan ng arterial na hadlang (atherosclerosis) at kolesterol sa dugo. Ang mga talakayan at aktibong pananaliksik na ito ay humantong sa paglikha ng mga epektibong gamot na nagpapababa ng kolesterol.
Noong 1913, ang N.N. Anichkov, isang kilalang siyentipiko at dalubhasa sa larangan ng eksperimentong patolohiya, ay naglathala ng isang gawain kung saan ipinakita niya na ang mga rabbits na pinapakain ng mayaman na may kolesterol ay nagkakaroon ng pinsala sa mga daluyan ng dugo na kahawig ng mga plato ng atherosclerotic sa mga daluyan ng mga matatandang tao. Isinasagawa ni Anichkov ang kanyang pananaliksik sa loob ng maraming mga dekada at nai-publish ang mga resulta sa kilalang mga journal sa Western. Sa kasamaang palad, ang kanyang data ay hindi naging batayan para sa isang modelo para sa pagbuo ng atherosclerosis sa mga tao, dahil sa oras na iyon ang hypothesis ay nananaig na ang sakit na ito ay isang natural na resulta ng pag-iipon at hindi mapipigilan. Gayunpaman, ang katibayan ay unti-unting naipon ng isang relasyon sa pagitan ng serum kolesterol at ang pag-unlad ng atherosclerosis (lipid hypothesis), at noong 1960s. ang ilang mga mananaliksik ay malinaw na nakasaad na ang sakit na ito ay maaaring gamutin ng mga gamot. Gayunpaman, ang kabaligtaran na punto ng pananaw ay umiiral hanggang sa nalathala noong 1984 ng mga resulta ng isang malawak na pag-aaral ng papel ng kolesterol na isinagawa ng US National Institute of Health (Coronary Primary Prevention Trial). Ang isang makabuluhang pagbaba ng istatistika sa dalas ng myocardial infarction at stroke na may pagbawas sa kolesterol ng dugo ay ipinakita. Sa pag-aaral na ito, ang kolesterol, isang resin ng anion exchange na nagbubuklod sa mga acid ng apdo, ay ginamit upang bawasan ang kolesterol. Ang mga resulta ay pinukaw ang paghahanap para sa mga bago, mas malakas na mga gamot na therapeutic. Dapat kong sabihin na sa pang-agham na mundo, ang mga pag-aalinlangan tungkol sa bisa ng lipid hypothesis ay ganap na nawala lamang sa pagdating ng mga statins sa huling bahagi ng 1980s - unang bahagi ng 1990s.
Ang unang statin ay natuklasan ni Akira Endo sa Sankyo sa Tokyo. Inilathala ni Endo ang kanyang gawain noong 1976, bagaman hinarap niya ang problema ng metabolismo ng kolesterol sa loob ng maraming taon. Noong 1971, iminungkahi niya na ang mga inhibitor ng kolesterol synthesis ay maaaring mapaloob din sa mga gumagawa ng kabute ng mga antibiotics na pinag-aralan sa oras na iyon. Sa loob ng maraming taon ng masinsinang gawain, sinuri niya ang higit sa 6,000 kultura ng iba't ibang mga kabute, hanggang sa siya ay dumating sa isang positibong resulta. Ang nagresultang compound ay tinawag na compactin. Ang sangkap na ito ay nagbaba ng kolesterol sa mga aso at unggoy. Ang mga pag-aaral na ito ay nakuha ng atensyon nina Michael Brown at Joseph Goldstein ng University of Texas Southwestern Medical School. Si Brown at Goldstein, kasama ang Endo, ay nagsimula ng isang pinagsamang pag-aaral at nakumpirma ang kanyang data. Ang mga pangunahing tagumpay ng unang mga klinikal na pagsubok ay kasangkot sa mga kumpanya ng parmasyutiko sa pagbuo ng mga bagong gamot. Sa Merck, isang pangkat na pinamunuan nina Alfred Alberts at Roy Wagelos ay naglunsad ng isang bagong screening ng mga kultura ng kabute at, bilang isang resulta ng pagsusuri ng isang kabuuang 18 kultura, natuklasan ang isa pang aktibong gamot. Ang bagong sangkap ay tinatawag na lovastatin. Gayunpaman, sa parehong oras, malawak na pinaniniwalaan na ang pangangasiwa ng mataas na dosis ng compactin sa mga aso ay humahantong sa pag-unlad ng kanser at ang paghahanap ng mga bagong statins noong 1980s. ay nasuspinde. Gayunpaman, sa oras na iyon, ang mga benepisyo ng paggamit ng mga statins upang gamutin ang mga pasyente na may familial hypercholesterolemia ay maliwanag na. Matapos ang maraming mga konsultasyon sa mga internasyonal na eksperto at ang Pagkain at Gamot na Pangangasiwa (FDA, USA), nagsimula ang pagbuo ng Merastatin. Malawak na pag-aaral sa susunod na dalawang dekada ay hindi inihayag ang carcinogenic na epekto ng lovastatin at ang bagong henerasyon ng mga gamot na lumitaw pagkatapos nito.
Fig. 1. Ang mga statins ay mga inhibitor ng NM G-CoA reductase. Paghahambing ng istraktura ng mevalonate at apat na produktong produktong parmasyutiko (statins) na pumipigil sa pagkilos ng NM G -CoA reductase.

Mga Statins pagbawalan ang pagkilos ng HMG - CoA - reductase, gayahin ang istraktura ng mevalonate, at sa gayon mai-block ang synthesis ng kolesterol. Sa mga pasyente na may hypercholesterolemia na sanhi ng isang kakulangan sa isang kopya ng LDL receptor gene, kapag kumukuha ng lovastatin, ang mga antas ng kolesterol ay nabawasan ng 30%. Ang gamot ay mas epektibo sa pagsasama sa mga espesyal na resin na nagbubuklod sa mga acid ng apdo at maiwasan ang kanilang reverse pagsipsip mula sa mga bituka.
Sa kasalukuyan, ang mga statins ay madalas na ginagamit upang babaan ang kolesterol ng plasma ng dugo. Kapag umiinom ng anumang gamot, ang tanong ay lumitaw tungkol sa kanilang hindi kanais-nais na mga epekto. Gayunpaman, sa kaso ng mga statins, maraming mga epekto, sa kaibahan, ay positibo. Ang mga gamot na ito ay maaaring mapukaw ang daloy ng dugo, ayusin na mayroon nang mga atherosclerotic plaques (upang hindi sila lumayo mula sa mga pader ng mga daluyan ng dugo at hindi makagambala sa daloy ng dugo), pagbawalan ang pagsasama ng platelet, at nagpapahina din ng mga nagpapaalab na proseso sa mga dingding ng mga daluyan ng dugo. Sa mga pasyente na kumukuha ng mga statins sa kauna-unahang pagkakataon, ang mga epektong ito ay nahayag kahit na bago magsimulang bumaba ang mga antas ng kolesterol, at posibleng nauugnay sa pagsugpo ng isoprenoid synthesis. Siyempre, hindi lahat ng epekto ng statins ay kapaki-pakinabang. Sa ilang mga pasyente (karaniwang kabilang sa mga kumukuha ng mga statins na pinagsama sa iba pang mga gamot na nagpapababa ng kolesterol), ang sakit ng kalamnan at kahinaan ng kalamnan ay maaaring mangyari, at kung minsan sa isang medyo malakas na anyo. Ang iba pang mga lubos na epekto ng statins ay nakarehistro din, na, sa kabutihang palad, bihirang mangyari. Sa karamihan ng mga pasyente, ang pagkuha ng mga statins ay maaaring pagbawalan ang pagbuo ng sakit sa cardiovascular. Tulad ng anumang iba pang gamot, ang mga statins ay dapat gamitin lamang bilang inirerekumenda ng iyong doktor.

Sa isang namamana na kawalan ng HDL kolesterol, ang mga antas ng kolesterol ay napakababa, na may sakit na Tangier, ang kolesterol ay praktikal na hindi tinutukoy. Ang parehong mga sakit sa genetic ay nagreresulta mula sa mga mutation sa ABC1 protein. Ang maliit na bahagi ng kolesterol ng HDL ay hindi maaaring makuha ang kolesterol mula sa mga selulang kulang sa ABC1, at ang mga cell na naubos sa kolesterol ay mabilis na tinanggal mula sa dugo at nawasak. Parehong ang namamana na kawalan ng HDL at Tangier disease ay napakabihirang (mas kaunti sa 100 mga pamilya na may sakit na Tangier ay kilala sa buong mundo), ngunit ang mga sakit na ito ay nagpapakita ng papel ng protina ng ABC1 sa pag-regulate ng mga antas ng plasma ng plasma. Dahil ang mababang antas ng HDL na antas ng HDL ay may ugnayan sa isang mataas na rate ng pinsala sa coronary artery, ang protina ng ABC1 ay maaaring maging kapaki-pakinabang na target para sa mga gamot na idinisenyo upang ayusin ang mga antas ng HDL. ■
Ang mga hormone ng Steroid ay nabuo sa pamamagitan ng paghahati ng side chain ng kolesterol at ang oksihenasyon nito.
Ang isang tao ay tumatanggap ng lahat ng kanyang mga hormone ng steroid mula sa kolesterol (Larawan 21-45). Dalawang klase ng mga hormone ng steroid ay synthesized sa adrenal cortex: mineral corticoids,na kinokontrol ang pagsipsip ng mga diorganikong mga ions (Na +, C l - at HC O 3 -) sa bato, at glucocorticoids, na tumutulong sa pag-regulate ng gluconeogenesis at bawasan ang nagpapasiklab na tugon. Ang mga sex hormones ay ginawa sa mga reproductive cell ng kalalakihan at kababaihan at sa inunan. Kabilang sa mga ito progesterone na kinokontrol ang babaeng reproductive cycle, androgens (hal. testosterone) at estrogen (estradiol), na nakakaapekto sa pag-unlad ng pangalawang sekswal na katangian sa kalalakihan at kababaihan, ayon sa pagkakabanggit. Ang mga hormone ng steroid ay may epekto sa napakababang konsentrasyon at samakatuwid ay synthesized sa medyo maliit na dami. Kung ikukumpara sa mga asing-gamot sa apdo, medyo maliit na kolesterol ang natupok para sa paggawa ng mga hormone ng steroid.

Fig. 21-45. Ang ilang mga steroid hormone ay nabuo mula sa kolesterol. Ang mga istruktura ng ilan sa mga compound na ito ay ipinapakita sa Fig. 10-19, v. 1.

Ang synthesis ng mga hormone ng steroid ay nangangailangan ng pag-alis ng marami o lahat ng mga carbon atoms sa "side chain" ng C-17 D-ring ng kolesterol. Ang pagtanggal ng chain sa gilid ay nangyayari sa mitochondria ng mga tisyu ng steroid na. Ang proseso ng pag-alis ay binubuo ng hydroxylation ng dalawang katabing mga atom ng carbon ng chain chain (C-20 at C-22), kung gayon ang pag-alis ng bono sa pagitan nila (Fig. 21-46). Ang pagbuo ng iba't ibang mga hormone ay may kasamang pagpapakilala ng mga atomo ng oxygen. Ang lahat ng mga reaksyon ng hydroxylation at oksihenasyon sa panahon ng biosynthesis ng steroid ay na-catalyzed ng mga oxidases na pinagsama-sama (magdagdag. 21-1) na gumagamit ng NА D PH, O 2 at mitochondrial cytochrome P-450.
Fig. 21-46. Ang cleavage ng side chain sa synthesis ng mga steroid hormone. Sa ganitong sistema ng oxidase na may isang halo-halong pag-andar na nag-oxidize sa mga katabing atom ng carbon, ang cytochrome P-450 ay kumikilos bilang isang carron elektron. Kasama rin sa proseso ay ang mga electron-transporting protein, adrenodoxin at adrenodoxin reductase. Ang sistemang ito ng side chain na paghahati ay natagpuan sa mitochondria ng adrenal cortex, kung saan nagaganap ang aktibong paggawa ng mga steroid. Ang Pregnenolone ay isang pangunguna sa lahat ng iba pang mga hormone ng steroid (Fig. 21-45).

Ang mga intermediate ng kolesterol na biosynthesis ay kasangkot sa maraming iba pang mga metabolic path.
Bilang karagdagan sa papel nito bilang isang intermediate ng biosynthesis ng kolesterol, ang isopentenyl pyrophosphate ay nagsisilbing isang aktibong paunang-natukoy sa synthesis ng isang malaking bilang ng mga biomolecules na nagsasagawa ng iba't ibang mga biological function (Fig. 21-47). Kabilang dito ang mga bitamina A, E at K, mga pigment ng halaman tulad ng karotina at ang chain ng chlorophyll phytol, natural na goma, maraming mahahalagang langis (halimbawa, ang mabangong base ng langis ng lemon, eucalyptus, musk), insekto juvenile hormone na nagreregula ng metamorphosis, dolichols, na maglingkod bilang lipid-soluble carriers sa kumplikadong synthesis ng polysaccharides, ubiquinone at plastoquinone - mga electron carriers sa mitochondria at chloroplast. Ang lahat ng mga molekulang ito ay isoprenoids sa istraktura. Mahigit sa 20,000 iba't ibang mga isoprenoid ay natagpuan sa likas na katangian, at daan-daang mga bago ang iniulat bawat taon.
Fig. 21-47. Ang pangkalahatang larawan ng biosynthesis ng isoprenoids. Ang mga istruktura ng karamihan sa mga produkto ng pagtatapos na ipinakita dito ay ibinibigay sa kap. 10 (v. 1).

Ang pagpapanggap (covalent attachment ng isang isoprenoid, tingnan ang Fig. 27-35) ay isang pangkaraniwang mekanismo na kung saan ang mga protina ay nakasalalay sa panloob na ibabaw ng mammalian cell lamad (tingnan ang Fig. 11-14). Sa ilang mga protina, ang nakatali na lipid ay kinakatawan ng isang 15-carbon farnesyl group, sa iba pa ito ay isang 20-carbon geranyl geranyl group. Ang dalawang uri ng lipid na ito ay naka-attach ng iba't ibang mga enzymes. Posible na ang mga reaksyon ng prenylation ay direktang protina sa iba't ibang mga lamad depende sa kung aling lipid ang nakalakip. Ang pagpapalabas ng protina ay isa pang mahalagang papel para sa mga isoprene derivatives - mga kalahok ng kolesterol metabolic pathway.
Buod ng Seksyon 21.4 Biosynthesis ng Cholesterol, Steroids, at Isoprenoids
■ Ang kolesterol ay nabuo mula sa acetyl-CoA sa isang kumplikadong pagkakasunud-sunod ng reaksyon sa pamamagitan ng mga tagapamagitan tulad ng β-hydroxy-β-methylglutaryl-CoA, mevalonate, dalawang aktibong isoprene dimethylallyl pyrophosphate at isopentenyl pyrophosphate. Ang kondensasyon ng mga yunit ng isoprene ay nagbibigay sa mga non-cyclic squalene, na kung saan ang mga siklista upang makabuo ng isang condensed ring system at steroid side chain.
■ Ang synthesis ng kolesterol ay nasa ilalim ng control ng hormonal at, bilang karagdagan, ay hinarang sa pamamagitan ng pagtaas ng konsentrasyon ng intracellular kolesterol, na nangyayari sa pamamagitan ng covalent modification at regulasyon ng transkrip.
■ Ang mga ester ng kolesterol at kolesterol ay dinadala ng dugo bilang mga lipoproteins ng plasma. Ang maliit na bahagi ng VLDL ay naglilipat ng kolesterol, kolesterol esters at triacylglycerol mula sa atay patungo sa iba pang mga tisyu, kung saan ang triacylglycerol ay na-clear ng lipoprotein lipase at ang VLDL ay na-convert sa LDL. Ang maliit na bahagi ng LDL na yaman sa kolesterol at kolesterol esters ay hindi direktang nakuha ng mga receptor sa pamamagitan ng endocytosis, habang ang apolipoprotein B-100 sa LDL ay kinikilala ng mga plasma receptor ng lamad. Tinatanggal ng HDL ang kolesterol sa dugo, inililipat ito sa atay. Ang mga kondisyon ng nutrisyon o mga depekto sa genetic sa metabolismo ng kolesterol ay maaaring humantong sa atherosclerosis at myocardial infarction.
■ Ang mga hormone na steroid (glucocorticoids, mineralocorticoids at sex hormones) ay nabuo mula sa kolesterol sa pamamagitan ng pagbabago ng chain chain at pagpapakilala ng mga atomo ng oxygen sa steroid system ng mga singsing. Maraming iba pang mga isoprenoid compound ay ginawa mula sa mevalonate sa pamamagitan ng paghalay ng isopentenyl pyrophosphate at dimethylallyl pyrophosphate kasama ang kolesterol.
■ Ang pagtatalo ng ilang mga protina ay nagtutuon sa kanila sa mga nagbubuklod na mga site na may mga lamad ng cell at mahalaga para sa kanilang biological na aktibidad.
Tanong 48. Ang regulasyon ng metabolismo ng mataas na fatty acid (β-oksihenasyon at biosynthesis). Sintesis ng malonyl CoA. Caretyylase ng Acetyl CoA, regulasyon ng aktibidad nito. Ang transportasyon ng acyl Co-a sa pamamagitan ng panloob na lamad ng mitochondria.
Pangunahing
ang halaga ng phenylalanine ay natupok
sa 2 paraan:
naka-on
sa mga squirrels,
lumiliko
sa tyrosine.
Ang pag-on
phenylalanine sa tyrosine pangunahin
kinakailangan upang alisin ang labis
phenylalanine, dahil ang mataas na konsentrasyon
Nakakalason sa mga cell. Edukasyon
Hindi talaga mahalaga ang tyrosine
mula sa kakulangan ng amino acid na ito
sa mga cell na halos hindi nangyayari.
Pangunahing
nagsisimula ang metabolismo ng phenylalanine
kasama ang hydroxylation nito (Fig. 9-29), sa
na nagreresulta sa tyrosine.
Ang reaksyon na ito ay catalyzed ng isang tiyak
m soaxy-nase - phenylalanine hydra (zsilase,
na nagsisilbing coferment
tetrahydrobiopterin (N4BP).
Ang aktibidad ng enzyme ay nakasalalay din
ang pagkakaroon ng Fe2.
Sa
ang atay ay pangunahing pinabilis na pagpapakilos
glycogen (tingnan ang seksyon 7). Gayunpaman stock
glycogen sa atay ay maubos
18-24 na oras ng pag-aayuno. Pangunahing mapagkukunan
asukal habang naubos ang mga stock
ang glycogen ay nagiging gluconeogenesis,
na nagsisimula upang mapabilis ang
Fig.
11-29. Mga pangunahing pagbabago sa metabolic
enerhiya kapag nagbabago sumisipsip
estado ng postabsorbent. CT
- mga katawan ng ketone, FA - fatty acid.
4-6 h
pagkatapos ng huling pagkain. Mga estruktura
Ang gliserol ay ginagamit para sa synthesis ng glucose,
amino acid at lactate. Sa mataas
rate ng synthesis ng glucose ng konsentrasyon
nabawasan ang mga fatty acid dahil sa
phosphorylation at hindi aktibo
acetyl CoA carboxylase at rate
pagtaas ng p-oksihenasyon. Gayunpaman,
nadagdagan ang supply ng taba sa atay
mga acid na inilipat
mula sa mga fat depot. Nabuo ang Acetyl-CoA
sa oksihenasyon ng mga fatty acid, ginagamit ito
sa atay para sa synthesis ng mga ketone na katawan.
Sa
adipose tissue na may pagtaas ng konsentrasyon
Binawasan ng glucoseagon ang rate ng synthesis
Ang TAG at lipolysis ay pinasigla. Stimulation
lipolysis - resulta ng pag-activate
hormon-sensitive TAG lipase
adipocytes sa ilalim ng impluwensya ng glucagon.
Ang Mga Fat Fat Acid ay Mahalaga
mapagkukunan ng enerhiya sa atay, kalamnan at
adipose tissue.
Kaya
sa gayon, sa panahon ng postabsorption
pinapanatili ang konsentrasyon ng glucose sa dugo
sa antas ng 80-100 mg / dl, at ang antas ng fat
ang mga asido at mga katawan ng ketone ay tumataas.
Asukal
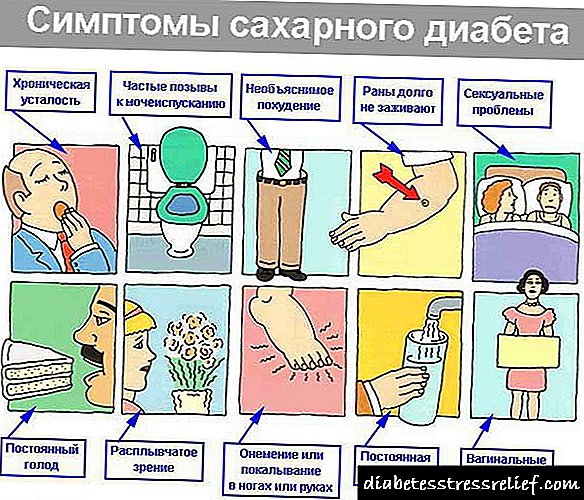
diabetes ay isang sakit na nangyayari
dahil sa ganap o kamag-anak
kakulangan sa insulin.
A.
Ang pangunahing klinikal na anyo ng asukal
diyabetis
Ayon kay
World Organization
diabetes sa pangangalagang pangkalusugan
inuri ayon sa pagkakaiba-iba
genetic factor at klinikal
dalawang pangunahing anyo: diabetes
Uri ng I - umaasa sa insulin (IDDM), at diyabetis
Uri ng II - independiyenteng non-insulin (NIDDM).
Regulasyon
synthesis ng zhk. Regulasyon ng enzyme
synthesis ng lcd - acetyl CoA carboxylase.
Ang enzim na ito ay kinokontrol ng maraming
mga paraan.
Aktibidad / Pagkakahiwalay
mga elemento ng subunit ng enzyme. Sa
hindi aktibong anyo ng acetyl CoA carboxylase
ay kumakatawan sa magkahiwalay na mga komplikado,
bawat isa ay binubuo ng 4 na subunits.
Ang activator ng enzyme ay citrate. Pinasisigla ito
kumbinasyon ng mga kumplikado, bilang isang resulta
kung saan tumataas ang aktibidad ng enzyme
. Inhibitor-palmitoyl-CoA. Tumawag siya
kumplikadong dissociation at pagbawas
aktibidad ng enzyme.
Phosphorylation / Dephosphorylation
acetyl CoA carboxylase. Sa
postabsorption estado o sa
gawaing pisikal na globo
adrenaline sa pamamagitan ng adenylate cyclase
ang sistema ay naisaaktibo ng prokinase A at
pasiglahin ang subunit phosphorylation
acetyl CoA carboxylase. Phosphorylated
ang enzyme ay hindi aktibo at ang synthesis ng mataba
huminto ang mga acid.
Sumisipsip
ang panahon ng insulin ay nag-activate ng phosphatase,
at acetyl-CoA carboxylase pumapasok
dephosphorylated na estado. Pagkatapos
sa ilalim ng impluwensya ng citrate ay nangyayari
polymerization ng mga protomer ng enzyme, at
nagiging aktibo siya. Bilang karagdagan sa pag-activate
enzyme, citrate ay gumaganap ng isa pa
gumana sa synthesis ng LCD. Sumisipsip
panahon sa mitochondria ng mga selula ng atay
nag-iipon ng citrate, kung saan
ang tirahan ng acyl ay dinadala sa
cytosol.
Regulasyon
rates-oksihenasyon rate.
Β-oksihenasyon-metabolic pathway,
matatag na naka-link sa gawain ng CPE at pangkalahatan
mga paraan ng catabolismo. Samakatuwid ang bilis nito
kinokontrol ng cell pangangailangan para sa
enerhiya i.e. sa pamamagitan ng mga ratio ng ATP / ADP at NADH / NAD, pati na rin ang reaksyon ng rate ng CPE at
karaniwang landas ng catabolism. Bilis
Ang β-oksihenasyon sa mga tisyu ay nakasalalay sa pagkakaroon
substrate, i.e.
sa dami ng mataba
mga asido na pumapasok sa mitochondria.
Libreng Fatty Acid Konsentrasyon
sa dugo ay tumataas sa pag-activate
lipolysis sa adipose tissue sa panahon ng pag-aayuno
sa ilalim ng impluwensya ng glucagon at sa panahon ng pisikal
gumana sa ilalim ng impluwensya ng adrenaline. Sa mga ito
ang mga fatty acid ay
nangingibabaw na mapagkukunan ng enerhiya
para sa mga kalamnan at atay, bilang isang resulta ng
Ang mga ox-oksihenasyon ay nabuo ng NADH at acetyl-CoA na pumipigil
pyruvate dehydrogenase complex.
Pagbabago ng pyruvate na bumubuo
mula sa glucose hanggang acetyl-CoA ay bumabagal.
Ang mga intermediate na metabolite ay makaipon
glycolysis at, sa partikular, glucose-6-phosphate.
Ang Glucose-6-phosphate ay pumipigil sa hexokinase
at samakatuwid ay humihina ang loob
ang paggamit ng glucose sa proseso
glikolisis. Samakatuwid, ang namamayani
paggamit ng lcd bilang pangunahing mapagkukunan
enerhiya sa kalamnan tissue at atay
nakakatipid ng glucose para sa nerve tissue at
pulang selula ng dugo.
Rate-oksihenasyon rate din
nakasalalay sa aktibidad ng enzyme
carnitine acyltransferases I.
Sa atay, ang enzyme na ito ay hinarang.
malonyl CoA, isang sangkap na nabuo
na may biosynthesis ng lcd. Sa panahon ng pagsisipsip
glikolisis ay isinaaktibo sa atay at
ang pagbuo ng acetyl-CoA ay nagdaragdag
mula sa pyruvate. Unang reaksyon ng synthesis
lcd conversion ng acetyl-CoA sa malonyl-CoA.
Pinipigilan ng Malonyl-CoA ang β-oksihenasyon ng lcd,
na maaaring magamit para sa synthesis
taba.
Edukasyon
malonyl-CoA mula sa acetyl-CoA-regulasyon
reaksyon sa biosynthesis lcd. Unang reaksyon
synthesis lcd conversion ng acetyl-CoA sa
malonyl CoA. Catalytic enzyme
reaksyon na ito (acetyl Coa carboxylase),
nabibilang sa klase ng mga ligases. Naglalaman siya
covalently bind na biotin. Sa una
co2 covalent reaksyon yugto
nagbubuklod sa biotin dahil sa enerhiya
Ang ATP, sa yugto 2 COO- inilipat
sa acetyl-CoA upang mabuo ang malonyl-CoA.
Acetyl CoA Carboxylase Enzyme Aktibidad
tinutukoy ang bilis ng lahat ng kasunod
mga reaksyon ng synthesis lc
ang citrate ay nag-activate ng isang enzyme sa cytosol
acetyl CoA carboxylase. Malonyl CoA sa
naman naman pinipigilan ang paglipat ng mas mataas
mataba acid mula sa cytosol hanggang matrix
pagbabawas ng mitochondria
panlabas na acetyl CoA: carnitine acyltransferase,
sa gayon pinapatay ang oksihenasyon ng mas mataas
mataba acids.
Acetyl-CoA Oxaloacetate →
HS-CoA Citrate

HSCOA ATP Citrate → Acetyl-CoA ADP Pi Oxaloacetate
Acetyl-CoA
sa cytoplasm ay nagsisilbing nagsisimula na substrate
para sa synthesis ng lcd, at oxaloacetate sa
Ang cytosol ay sumasailalim sa mga pagbabagong-anyo sa
ang resulta ng kung saan ang pyruvate ay nabuo.
Biosynthesis ng kolesterol
Ang biosynthesis ng kolesterol ay nangyayari sa endoplasmic reticulum. Ang mapagkukunan ng lahat ng mga carbon atoms sa molekula ay acetyl-SCoA, na nagmumula rito mula sa mitochondria sa citrate, tulad ng synthesis ng mga fatty acid. Kinokonsumo ng kolesterol biosynthesis ang 18 ATP molekula at 13 NADPH molekula.
Ang pagbuo ng kolesterol ay nangyayari sa higit sa 30 reaksyon, na maaaring maipangkat sa maraming yugto.
1. Sintesis ng mevalonic acid.
Ang unang dalawang reaksiyon ng synthesis ay nag-tutugma sa mga reaksyon ng ketogenesis, ngunit pagkatapos ng synthesis ng 3-hydroxy-3-methylglutaryl-ScoA, pumapasok ang enzyme. hydroxymethyl-glutaryl-ScoA reductase (HMG-SCOA reductase), na bumubuo ng mevalonic acid.
 |